Resumos
Avaliou-se a taxa de assimilação de CO2 e o potencial da água nas folhas, em laranjeiras `Valência' com 18 meses de idade, em vasos, enxertadas em quatro tipos de porta-enxertos: limoeiro `Cravo', Trifoliata, citrange `Troyer' e tangerina `Cleópatra'. A deficiência hídrica foi obtida pela suspensão da irrigação até que a taxa de assimilação de CO2 atingisse valores aproximadamente nulos, quando, então, foram reirrigadas. Avaliou-se, também, a condutividade hidráulica dos porta-enxertos utilizados. Sem deficiência hídrica, a taxa de assimilação de CO2 foi menor nas plantas sobre os porta-enxertos `Troyer' e `Cleópatra', quando comparadas àquelas plantas sobre limoeiro `Cravo', que não se diferenciaram do Trifoliata. As maiores taxas de assimilação de CO2 foram observadas por volta das 11h, com 8 mmol/m2.s, seguidas por uma diminuição, provavelmente, em vista do aumento do déficit de pressão de vapor e da temperatura ambiente entre as 11 e as 14h. Após a suspensão da irrigação, a queda do potencial da água na folha foi semelhante em todas as plantas, independentemente dos porta-enxertos usados, porém, sobre `Cravo', a taxa de assimilação de CO2 decaiu mais rapidamente e recuperou-se mais vagarosamente. As plantas sobre esse porta-enxerto apresentaram, durante todo o período de deficiência hídrica, menor taxa de assimilação de CO2 que aquelas sobre Trifoliata e `Cleópatra', e semelhante ao `Troyer'. As plantas sobre Trifoliata recuperaram-se em três dias, enquanto as outras não se recuperaram totalmente. A condutividade hidráulica foi superior para o limoeiro `Cravo' e semelhante entre os demais porta-enxertos.
Citrus sinensis; condutividade hidráulica das raízes; potencial da água; taxa de assimilação líquida de CO2
The CO2 assimilation rate and leaf water potential were analyzed in 18-months-old `Valencia' orange tree, in pots. The plants were grafted on four different rootstocks: `Rangpur' lime, Trifoliata, `Troyer' citrange and `Cleopatra' mandarin. Plants were submitted to water stress until the CO2 assimilation rate was negligible when they were rehydrated. It was also analysed the hydraulic conductivity of the rootstocks used. Under well watered conditions, CO2 assimilation rate was lower in plants grafted on `Troyer' and `Cleopatra' when compared with plants grafted on `Cravo' which was not different from plants grafted on Trifoliata. Throughout the day the greatest CO2 assimilation rates were observed by 9:00h, with 8mmol/m2.s, followed by a decrease due possibly to the increase in the vapour water deficit and room temperature, from 11:00h through 14:00h. After withholding water, in the decreasing at the leaf water potential was similar to all the plants, regardless of the rootstocks used. Nevertheless, CO2 assimilation rate decreased more rapidly, after withholding water, and it was recovered more slowly after rehydration, in plants grafted on `Cravo'. These plants also presented, during the entire water deficit period, lower CO2 assimilation rate than plants grafted on Trifoliata and `Cleopatra', and similar to `Troyer'. Plants grafted on Trifoliata recovered CO2 assimilation rate in 3 days while those grafted on other rootstocks did not recover entirely in that period. The hydraulic conductivity was higher in `Cravo' lemon tree and there were no differences among the values found in the other rootstocks.
Citrus sinensis; root hydraulic conductivity; water potential; net CO2 assimilation rate
I. BOTÂNICA, FITOQUíMICA E FISIOLOGIA DE PLANTAS
FOTOSSÍNTESE DE LARANJEIRA `VALÊNCIA' ENXERTADA SOBRE QUATRO PORTA-ENXERTOS E SUBMETIDA À DEFICIÊNCIA HÍDRICA(1) (1) Parte da Tese de Mestrado do primeiro autor em Fisiologia e Bioquímica de Plantas. Projeto financiado pela FAPESP. Recebido para publicação em 2 de julho de 1997 e aceito em 31 de janeiro de 1998.
CAMILO LÁZARO MEDINA(2) (2) Pós-graduando em Fisiologia e Bioquímica de Plantas da Escola Superior de Agricultura Luiz de Queiroz, ESALQ-USP, Piracicaba (SP). Centro de Ecofisiologia e Biofísica, Instituto Agronômico (IAC), Caixa Postal 28, 13001-970 Campinas (SP). (3) Centro de Ecofisiologia e Biofísica, IAC. (4) Pós-graduando em Irrigação e Drenagem, ESALQ-USP CPATSA-EMBRAPA, Caixa Postal 23, 56300-000 Petrolina (PE). (5) Com bolsa de produtividade em pesquisa do CNPq. , EDUARDO CARUSO MACHADO(3,5) (2) Pós-graduando em Fisiologia e Bioquímica de Plantas da Escola Superior de Agricultura Luiz de Queiroz, ESALQ-USP, Piracicaba (SP). Centro de Ecofisiologia e Biofísica, Instituto Agronômico (IAC), Caixa Postal 28, 13001-970 Campinas (SP). (3) Centro de Ecofisiologia e Biofísica, IAC. (4) Pós-graduando em Irrigação e Drenagem, ESALQ-USP CPATSA-EMBRAPA, Caixa Postal 23, 56300-000 Petrolina (PE). (5) Com bolsa de produtividade em pesquisa do CNPq. e JOSÉ MARIA PINTO (4) (2) Pós-graduando em Fisiologia e Bioquímica de Plantas da Escola Superior de Agricultura Luiz de Queiroz, ESALQ-USP, Piracicaba (SP). Centro de Ecofisiologia e Biofísica, Instituto Agronômico (IAC), Caixa Postal 28, 13001-970 Campinas (SP). (3) Centro de Ecofisiologia e Biofísica, IAC. (4) Pós-graduando em Irrigação e Drenagem, ESALQ-USP CPATSA-EMBRAPA, Caixa Postal 23, 56300-000 Petrolina (PE). (5) Com bolsa de produtividade em pesquisa do CNPq.
RESUMO
Avaliou-se a taxa de assimilação de CO2 e o potencial da água nas folhas, em laranjeiras `Valência' com 18 meses de idade, em vasos, enxertadas em quatro tipos de porta-enxertos: limoeiro `Cravo', Trifoliata, citrange `Troyer' e tangerina `Cleópatra'. A deficiência hídrica foi obtida pela suspensão da irrigação até que a taxa de assimilação de CO2 atingisse valores aproximadamente nulos, quando, então, foram reirrigadas. Avaliou-se, também, a condutividade hidráulica dos porta-enxertos utilizados. Sem deficiência hídrica, a taxa de assimilação de CO2 foi menor nas plantas sobre os porta-enxertos `Troyer' e `Cleópatra', quando comparadas àquelas plantas sobre limoeiro `Cravo', que não se diferenciaram do Trifoliata. As maiores taxas de assimilação de CO2 foram observadas por volta das 11h, com 8 mmol/m2.s, seguidas por uma diminuição, provavelmente, em vista do aumento do déficit de pressão de vapor e da temperatura ambiente entre as 11 e as 14h. Após a suspensão da irrigação, a queda do potencial da água na folha foi semelhante em todas as plantas, independentemente dos porta-enxertos usados, porém, sobre `Cravo', a taxa de assimilação de CO2 decaiu mais rapidamente e recuperou-se mais vagarosamente. As plantas sobre esse porta-enxerto apresentaram, durante todo o período de deficiência hídrica, menor taxa de assimilação de CO2 que aquelas sobre Trifoliata e `Cleópatra', e semelhante ao `Troyer'. As plantas sobre Trifoliata recuperaram-se em três dias, enquanto as outras não se recuperaram totalmente. A condutividade hidráulica foi superior para o limoeiro `Cravo' e semelhante entre os demais porta-enxertos.
Termos de indexação: Citrus sinensis, condutividade hidráulica das raízes, potencial da água, taxa de assimilação líquida de CO2.
ABSTRACT
PHOTOSYNTHESIS OF `VALENCIA' ORANGE TREE GRAFTED ON FOUR ROOTSTOCKS AND SUBMITTED TO WATER DEFICIT
The CO2 assimilation rate and leaf water potential were analyzed in 18-months-old `Valencia' orange tree, in pots. The plants were grafted on four different rootstocks: `Rangpur' lime, Trifoliata, `Troyer' citrange and `Cleopatra' mandarin. Plants were submitted to water stress until the CO2 assimilation rate was negligible when they were rehydrated. It was also analysed the hydraulic conductivity of the rootstocks used. Under well watered conditions, CO2 assimilation rate was lower in plants grafted on `Troyer' and `Cleopatra' when compared with plants grafted on `Cravo' which was not different from plants grafted on Trifoliata. Throughout the day the greatest CO2 assimilation rates were observed by 9:00h, with 8mmol/m2.s, followed by a decrease due possibly to the increase in the vapour water deficit and room temperature, from 11:00h through 14:00h. After withholding water, in the decreasing at the leaf water potential was similar to all the plants, regardless of the rootstocks used. Nevertheless, CO2 assimilation rate decreased more rapidly, after withholding water, and it was recovered more slowly after rehydration, in plants grafted on `Cravo'. These plants also presented, during the entire water deficit period, lower CO2 assimilation rate than plants grafted on Trifoliata and `Cleopatra', and similar to `Troyer'. Plants grafted on Trifoliata recovered CO2 assimilation rate in 3 days while those grafted on other rootstocks did not recover entirely in that period. The hydraulic conductivity was higher in `Cravo' lemon tree and there were no differences among the values found in the other rootstocks.
Index terms: Citrus sinensis, root hydraulic conductivity, water potential, net CO2 assimilation rate.
1. INTRODUÇÃO
Na enxertia - o método mais utilizado para a formação de mudas cítricas - os porta-enxertos empregados na cultura afetam várias características, como: produtividade (Wutscher, 1979; Atkinson, 1980; Wutscher & Bistline, 1988; Pompeu Jr., 1991); desenvolvimento vegetativo (Teófilo Sobrinho, 1991); tolerância a doenças (Guirado et al., 1991); qualidade interna e externa dos frutos (Wardowski et al., 1986); relações hídricas e trocas gasosas (Camacho et al., 1974; Crocker et al., 1974; Syvertsen & Graham, 1985, Yelenosky, 1991) e a adaptabilidade à hipoxia e à salinidade (Lloyd et al.,1987).
Apesar de vários porta-enxertos disponíveis, a citricultura brasileira está quase totalmente apoiada sobre o limoeiro `Cravo' (Pompeu Jr., 1991). Essa situação é inconveniente, como se observou na década dos 40s, quando o vírus da tristeza dizimou os plantios de laranja-doce enxertados sobre laranjeira `Azeda' (Kimati & Galli, 1980). Além disso, uma combinação mais adequada de porta-enxerto e copa deve ser adaptada às condições em que o pomar será implantado.
No Brasil, apesar da incidência de períodos de déficits hídricos temporários em várias regiões citrícolas, predomina o cultivo sem irrigação (Ortolani et al., 1991). Sob deficiência hídrica, ocorre o fechamento dos estômatos, diminuindo a difusão do vapor d'água para a atmosfera e a difusão do CO2 para o mesofilo foliar, com depressão da fotossíntese e da produtividade. Diferentes combinações de porta-enxerto e copa revelam comportamentos característicos em relação às trocas gasosas e à hídrica (Castle et al., 1989), afetando o grau de tolerância à seca. Isso, contudo, pode estar relacionado a características do sistema radicular de cada porta-enxerto utilizado, como profundidade, arquitetura e condutividade hidráulica das raízes (Kriedemann & Barrs, 1981; Syvertsen & Graham, 1985; Hale & Orcutt, 1987).
O objetivo deste trabalho foi avaliar a influência dos porta-enxertos citrange `Troyer', tangerina `Cleópatra', limoeiro `Cravo' e Trifoliata, nas trocas gasosas e nas relações hídricas da laranjeira `Valência' quando submetida à deficiência hídrica.
2. MATERIAL E MÉTODOS
Mudas de laranjeira `Valência' (Citrus sinensis Osbeck) com 18 meses de idade, enxertadas sobre os porta-enxertos citrange `Troyer', tangerina `Cleópatra' (Citrus reshni Hort. ex Tan.), limoeiro `Cravo' (Citrus limonia Osbeck) e Poncirus trifoliata (Trifoliata), foram transplantadas para vasos com 0,45 m de altura por 0,18 m de diâmetro, com substrato composto de três partes de terra e uma de areia, sobre uma camada de 0,02 m de pedras britadas. O substrato apresentou 38 mg/dm3 de P em extrator de resina; 40 mmol/dm3 de Ca; 11 mmol/dm3 de Mg; 18 mmol/dm3 de H + Al; 0,1 mmol/dm3 de K; 1,3% de matéria orgânica; pH em solução CaCl2 de 5,8. Adubações via solo foram feitas quinzenalmente, totalizando 4,46 g de N; 0,54 g de P; 2,10 g de K; 0,35 g de S; 100 mg de Zn; 50 mg de Mg; 15 mg de B e 30 mg de Cu, por vaso, durante os cinco meses de desenvolvimento da planta. Também, foram efetuadas cinco pulverizações foliares com solução de 0,15% de ZnSO4, 0,15% de MnSO4 com 0,5% de KNO3. Sete dias antes do início das medições, as plantas foram irrigadas com um litro de solução nutritiva de Hoagland & Arnon (1950).
Cultivaram-se laranjeiras em casa de vegetação e dispostas em linhas distantes de 1,80 m por 0,65 m entre plantas, no sentido norte-sul, em quatro blocos (repetições) com oito vasos em cada um, sendo dois vasos para cada porta-enxerto. Por ocasião das medidas, as raízes das plantas encontravam-se espalhadas por todo o volume dos vasos. Até o início dos tratamentos, as plantas foram irrigadas diariamente. A deficiência hídrica foi induzida em um dos dois vasos de cada variedade de porta-enxerto, por bloco, enquanto o outro recebeu água suficiente para não desenvolver deficiência hídrica.
A quantidade média diária de água adicionada em cada vaso foi estimada da seguinte forma: saturou-se o substrato no vaso e permitiu-se a drenagem por 12 horas, à noite. A seguir, mediu-se a massa úmida na capacidade de campo e após 24 horas. A quantidade de água consumida foi totalmente reposta no tratamento sem deficiência e, naquele com deficiência hídrica, nos primeiros sete dias foi reposta somente 50% da água consumida e, entre o oitavo e o décimo terceiro dia, suspendeu-se totalmente a irrigação. Dessa forma, a deficiência hídrica foi imposta de modo vagaroso e crescente entre os dias 14 e 23 de janeiro, quando as plantas foram reirrigadas. Diariamente, determinou-se o potencial da água nas folhas (Y), utilizando-se uma câmara de pressão com a técnica descrita por Kaufmann (1968). As medidas foram feitas às 6 (Y6) e às 13h (Y13), com três repetições por tratamento. A umidade do substrato foi determinada à profundidade de 0,15 m em relação à superfície do vaso. Amostras de substrato foram retiradas diariamente, determinando-se a massa fresca e, posteriormente, a seca, após secagem em estufa por 24 horas a 100 oC.
As taxas de assimilação líquida de CO2 (A) e a densidade de fluxo de fótons fotossinteticamente ativos (DFFFA) foram medidas com um analisador portátil de fotossíntese por radiação infravermelha, Li-6200 (LICOR Inc., Lincoln, USA), com técnica descrita por Vu et al. (1986). As medidas foram feitas em condições naturais, em folhas com seis meses de idade, totalmente expandidas, dispostas em posições semelhantes em relação à planta e à incidência de radiação solar, em intervalos de duas horas no período compreendido entre as 8 e as 16h. As plantas deficientes em água, após terem atingido um valor de assimilação de CO2 próximo de zero, foram irrigadas novamente, repetindo-se as mesmas medidas do período de recuperação.
Para a estimativa da condutividade hidráulica do sistema radicular, semearam-se 98 sementes de cada porta-enxerto, em tubetes com 0,125 m de altura com substrato estéril composto de vermiculita e matéria orgânica fertilizada (Plantimax/Eucatex). Adubou-se mediante solução nutritiva de Hoagland & Arnon (1950), em irrigações diárias totalizando 500 ml da solução por plântula. Efetuaram-se as medidas de condutividade hidráulica quando as plântulas atingiram altura de 0,2 m e diâmetro do caule de 0,004 m.
A condutividade hidráulica das raízes foi medida com cinco repetições por porta-enxerto, pelo método descrito por Graham & Syvertsen (1984). As plântulas foram transferidas para o laboratório, onde permaneceram por doze horas sob baixa iluminação (10 mmol/m2.s) a 26 oC. Nesse período, os tubetes ficaram mergulhados em água destilada à altura de 0,05 m, até o momento das medidas, quando o caule da plântula foi cortado ao redor de 0,05 m acima do colo da planta. O tubete, com sistema radicular intacto e substrato saturado, foi colocado dentro de uma câmara de pressão e a base da plântula, mergulhada parcialmente em água, à temperatura de 25 o C. A pressão foi gradativamente aumentada até 0,5 MPa. Após o equilíbrio inicial (dez minutos), determinou-se a quantidade de líquido exsudado em intervalos de dois minutos. Após a determinação do comprimento das raízes pelo método de interseção de linhas (Tennant, 1975), a condutividade hidráulica foi expressa como massa de exsudato em mg/m.s.MPa.
A análise estatística foi em blocos ao acaso e a comparação de médias, pelo teste de Duncan a 5%.
3. RESULTADOS E DISCUSSÃO
3.1 Plantas sem deficiência hídrica
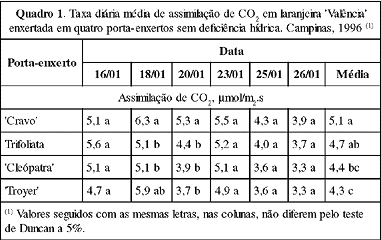
Os valores médios da taxa de assimilação de CO2 das laranjeiras 'Valência' enxertadas em citrange `Troyer' e tangerina `Cleópatra' foram menores que os do limoeiro `Cravo'. `Troyer' também induziu menor taxa de assimilação de CO2 que Trifoliata e este, por sua vez, não se diferenciou do `Cravo' e da `Cleópatra' (Quadro 1).
Tais diferenças mostram que os porta-enxertos afetaram as trocas gasosas, conforme também relatado por outros autores (Camacho et al., 1974; Crocker et al., 1974; Syvertsen & Grahan, 1985; Yelenosky, 1991). Sinclair & Allen (1982) verificaram que porta-enxertos com maior condutividade hidráulica podem manter as folhas mais hidratadas de dia, tendo uma relação direta com a abertura estomática e, por conseguinte, com as trocas gasosas.
Analogamente, o limoeiro `Cravo', que apresentou maior condutividade hidráulica (Quadro 2), pode ter induzido maior assimilação de CO2.
O Trifoliata é um porta-enxerto que induz menor desenvolvimento das copas (Teófilo Sobrinho, 1991). Essa característica, porém, parece não estar relacionada à taxa de assimilação de CO2, já que apresentou valor semelhante ao `Cravo'.
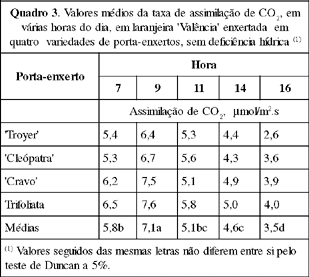
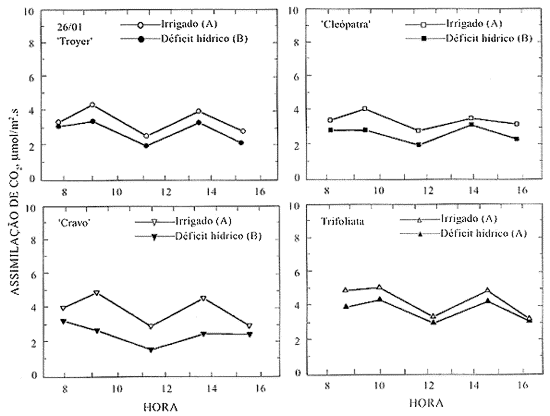
Os maiores valores de taxa de assimilação de CO2, próximos a 8 mmol/m2.s, foram observados entre as 9 e as 11 horas, quando a DFFFA esteve acima de 700 mmol/m2.s(Figuras 2 a 6 e Quadro 3). Esses valores de assimilação de CO2 estão na faixa dos encontrados para os citros (Syvertsen, 1984). Apesar de a DFFFA para a saturação lumínica dos citros situar-se ao redor de 700 mmol/m2.s (Syvertsen, 1984), houve diminuição da taxa de assimilação de CO2 após as 11 h, embora a ocorrência de valores maiores de radiação solar (Quadro 4). Essa queda na taxa de assimilação de CO2 pode estar associada ao aumento do déficit de pressão de vapor de água no ar (DPV), que, freqüentemente, ocorre com o aumento de temperatura, nas horas de maior intensidade de radiação solar, como foi observado por Khairi & Hall (1976); Levy (1980); Sinclair & Allen (1982).
Sinclair & Allen (1982) observaram queda na assimilação de CO2 quando o DPV foi maior que 1,52 kPa. Esses valores são menores que os atingidos neste experimento (Quadro 4). Visto que a faixa de temperatura ideal para a atividade fotossintética dos citros está entre 25 e 31 oC e, praticamente, cessa entre 38 e 40 oC (Reuther, 1977), as temperaturas acima de 31 oC, que ocorreram após as 10 h, também devem ter contribuído para diminuir a assimilação de CO2 (Quadro 4).
Segundo Hale & Orcutt (1987), temperaturas altas aumentam a fotorrespiração, contribuindo também para diminuir a taxa de assimilação de CO2.
3.2 Plantas sob deficiência hídrica
A queda no teor de umidade do substrato das plantas submetidas à deficiência hídrica foi semelhante para todos os tratamentos, independentemente do porta-enxerto usado, isto é, de 22% no início do tratamento até ao redor de 12% no final (Figura 1).
Umidade do substrato em vasos com laranjeira 'Valência' enxertada em quatro variedades de porta-enxertos.
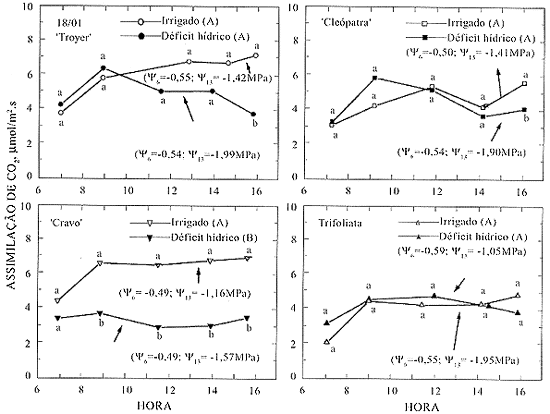
Dois dias após a suspensão da irrigação, apesar de já ter ocorrido um declínio significativo na umidade do solo (Figura 1), não se observaram diferenças significativas nos potenciais da água e nas taxas de assimilação de CO2 entre os tratamentos irrigados e sob deficiência hídrica (Figura 2).
Taxa de assimilação de CO2 em laranjeira 'Valência' sobre os quatro porta-enxertos, dois dias após a suspensão da irrigação. Tratamentos irrigados e seus correspondentes com déficit hídrico, seguidos da mesma letra, não diferem entre si pelo teste de Duncan a 5%.
Sob deficiência hídrica, no quinto e no sétimo dia, somente o limoeiro `Cravo' apresentou menor taxa de assimilação de CO2 em relação à testemunha (Figuras 3 e 4). Essa queda se iniciou quando Y6 e Y13 atingiram os valores de, respectivamente, -0,54 e -1,57 MPa, enquanto a testemunha apresentou -0,49 e -1,16 MPa. Para os outros tratamentos, também se verificou redução no potencial da água. Contudo, não ocorreu decréscimo significativo na absorção diária de CO2. Assim, o tratamento 'Cravo' apresentou queda mais acentuada na assimilação de CO2 em valores de Y6 e Y13 semelhantes aos dos outros tratamentos (Figuras 3 e 4).
Taxa de assimilação de CO2 em laranjeira 'Valência' sobre os quatro porta-enxertos, quatro dias após a suspensão da irrigação. Tratamentos irrigados e o correspondente com deficiência hídrica, seguidos com a mesma letra maiúscula, indicam que as taxas de assimilação média diária não diferem entre si pelo teste de Duncan a 5%. Valores pontuais seguidos das mesmas letras minúsculas não diferem entre si pelo teste de Duncan a 5%.
Taxa de assimilação de CO2 em laranjeira 'Valência' sobre os quatro porta-enxertos, seis dias após a suspensão da irrigação. (Ver legenda da Figura 3.)
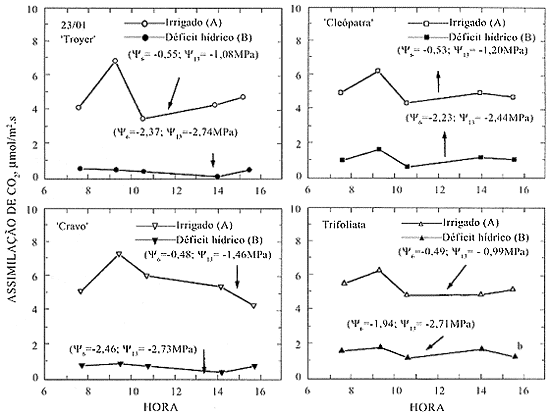
No décimo dia de deficiência hídrica, a taxa de assimilação de CO2 reduziu acentuadamente, atingindo valores próximos a zero, independentemente do porta-enxerto. O potencial da água, Y6 e Y13, foi reduzida e as folhas ficaram murchas durante o dia todo, mostrando claros sinais de estresse (Figura 5).
Taxa de assimilação de CO2 em laranjeira 'Valência' sobre os quatro porta-enxertos, dez dias após a suspensão da irrigação. Tratamentos irrigados e seus correspondentes com déficit hídrico, seguidos da mesma letra, não diferem entre si pelo teste de Duncan a 5%.
Após a irrigação, no dia 23, as plantas recuperaram o potencial da água no dia seguinte (dados não apresentados), porém a taxa de assimilação de CO2 não se recuperou mesmo após dois dias (Figura 6 ). A recuperação da assimilação foi menor para o tratamento `Cravo' (44,8%), sendo, para `Troyer', `Cleópatra' e Trifoliata, próxima de 57% em relação às testemunhas.
Figura 6. Taxa de assimilação de CO2 em laranjeira 'Valência' sobre os quatro porta-enxertos, dois dias após a reirrigação. Tratamentos irrigados e seus correspondentes com déficit hídrico, seguidos da mesma letra, não diferem entre si pelo teste de Duncan a 5%.
No terceiro dia de recuperação, somente as plantas sobre Trifoliata apresentaram taxa de assimilação de CO2 estatisticamente semelhante à da testemunha irrigada. As taxas de assimilação de CO2 das plantas sobre os outros porta-enxertos recuperaram-se parcialmente em relação aos respectivos controles, ou seja, 64% no `Cravo' e cerca de 79% no `Troyer' e `Cleópatra' (Figura 7). Apesar de as plantas sobre `Cravo' terem sua taxa de assimilação de CO2 reduzida um dia mais cedo (Figura 3) que as plantas sobre os outros porta-enxertos, isso não implicou manutenção de maior potencial da água na folha, nem em recuperação mais rápida.
Taxa de assimilação de CO2 em laranjeira 'Valência' sobre os quatro porta-enxertos, três dias após a reirrigação. Tratamentos irrigados e seus correspondentes com déficit hídrico, seguidos da mesma letra, não diferem entre si pelo teste de Duncan a 5%.
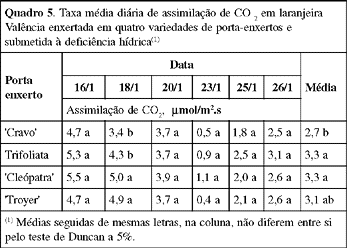
Na análise global de todos os dados relativos a plantas submetidas à deficiência hídrica, a taxa de assimilação média de CO2, no período experimental, foi menor para o `Cravo' em relação à da Trifoliata e à da `Cleópatra' (Quadro 5).
Plantas enxertadas sobre limoeiro `Cravo' apresentam, em condições de campo, maior tolerância à seca que as enxertadas sobre os outros porta-enxertos aqui utilizados (Pompeu Jr., 1991). Esses resultados sugerem, inicialmente, que a maior tolerância induzida pelo limoeiro `Cravo', no campo, pode não estar relacionada com características transmitidas pelo porta-enxerto à parte aérea e, sim, com características relacionadas ao sistema radicular do próprio porta-enxerto que, em condições de vaso, não se manifestam. Kriedemann & Barrs (1981) relatam que porta-enxertos de limões e limas favorecem as plantas nas condições de deficiência hídrica, pela maior profundidade e ramificação do sistema radicular. Em relação a este, deve-se considerar, além do hábito de crescimento (vigor e distribuição das raízes), a condutividade hidráulica das raízes, sua capacidade de produção de hormônios que afetariam o comportamento do próprio crescimento das raízes e o funcionamento dos estômatos (Stevens & Westwood, 1984; Syvertsen & Graham, 1985; Turner et al., 1985).
A maior condutividade hidráulica para o porta-enxerto limoeiro `Cravo', embora possa auxiliar a manutenção do potencial da água, por conferir menor resistência ao transporte de água para as folhas (Kaufmann, 1968, Sinclair & Allen, 1982), não impediu que ocorresse diminuição da taxa de assimilação de CO2 quando as plantas estiveram submetidas ao estresse. A diminuição da assimilação de CO2, na fase de desenvolvimento da deficiência hídrica, pode estar relacionada à diminuição da condutância estomática, aumento da resistência ao transporte interno de CO2 e danos por deficiência hídrica ao aparelho fotossintético (Berry & Downton, 1982). Desde que o estresse hídrico por si pode reduzir a condutividade da raiz (Ramos & Kaufmann, 1976, Levy et al., 1983), a deficiência hídrica mais acentuada no limoeiro `Cravo' pode também ter retardado sua recuperação.
Embora o comportamento possa ser alterado em condições de campo, a recuperação mais rápida para o Trifoliata nas condições estudadas sugere que esse porta-enxerto induziu maior tolerância à seca às copas. O Trifoliata adapta-se bem em solos que dificultam o desenvolvimento do sistema radicular, como os mais argilosos (Pompeu Jr., 1991), que, aparentemente, manifestaram-se nas condições de limitação ao desenvolvimento das raízes, conforme aquelas presentes aqui em estudo.
YELENOSKY, G. Response and adaptation of citrus trees to environmental stresses. Israel J. Bot. 40:239-250, 1991.
- ATKINSON, D. The distribution and effectiveness of the roots of tree crops. In: JANICK, J., ed. Horticultural Reviews II, Westport, AVI, 1980. p.429-490.
- BERRY, J.A. & DOWNTON, W.J.S. Environmental regulation of photosynthesis. In GOVINDJEE, E. Photosynthesis: development, carbon metabolism, and plant production. New York, Academic Press, 1982. v.2, p.306-308.
- CAMACHO, S.E.; KAUFMANN, M.R. & HALL, A.E. Leaf water potential response to transpiration by citrus. Physiologia Plantarum, Copenhagen, 31:101-105, 1974.
- CASTLE W.S; TUCKER, D.P.H.; KREZDORN, A.H. & YOUTSEY, C.O. Rootstocks for Florida Citrus Gainesville, Institute of Food and Agricultural Science, University of Florida, 1989. 47p.
- CROCKER, T.E.; BELL, W.D. & BARTHOLIC, J.F. Scholander pressure bomb technique to assess the relative leaf water of 'Orlando' tangelo scion as influenced by varius citrus rootstocks. HortScience, Alexandria, 9:453-455, 1974.
- GRAHAN, J.H. & SYVERSTSEN, J.P. Influence of VAM on hydraulic conductivity of roots of two citrus rootstocks. New Phytologist, Oxford, 97: 277-284, 1984.
- GUIRADO, N.; MULLER, G.W. & PRATES, H. S. Declínio dos citros. In: RODRIGUES, O.: VIÉGAS, F.; POMPEU JUNIOR., J. & AMARO, A.A., eds. Citricultura brasileira 2.ed. Campinas, Fundaçăo Cargill, 1991. p. 722-734.
- HALE, M.G. & ORCUTT, D.M. The physiology of plants under stress New York, John Willey, 1978. p.26-43.
- HOAGLAND, D.R. & ARNON, D.I. The water-culture method for growing plants without soil Berkeley, California Agricultural Experiment Station, 1950. 32p. (Circular, 347.)
- KAUFMANN, M. R. Evaluation of the pressure chamber method for measurement of water stress in citrus. Proceedings of American Society of Horticultural Science, New York, 93:186-198, 1968.
- KHAIRI, M.M.A. & HALL, A.E. Temperature and humidity effects on net photosyntesis and transpiration of citrus. Physiologia Plantarum, Copenhagen, 36:29-34, 1976.
- KIMATI, H. & GALLI, F.E. Doenças dos citros. In: GALLI, F.E., ed. Manual de fitopatologia Săo Paulo, Agronômica Ceres, 1980. p. 213-235.
- KRIEDEMANN, P. E. & BARRS, H.D. Citrus orchards. In: KOZLOWSKI, T.T. Water deficits and plant growth VI. Woody plant communities. New York, Academic Press, 1981. p. 325-418.
- LEVY, Y. Effect of evaporative demand on water relations of Citrus limon. Annals of Botany, London, 46:695-700, 1980.
- LEVY, Y.; SYVERTSEN, J.P. & NEMEC, S. Effect of drought stress ad VAM on citrus transpiration and hydraulic conductivity of roots. New Phytologist, Oxford, 93:61-66, 1983.
- LLOYD, J.; KRIEDEMANN, P.E. & SYVERTSEN, J.P. Gas exchange, water relations and ion concentration of leaves on salt stressed `Valęncia' orange, Citrus sinensis (L.) Osbeck. Australian Journal of Plant Physiolog, Melbourn, 14:387-396, 1987.
- ORTOLANI, A.A.; PEDRO JUNIOR, M.J. & ALFONSI, R.R. Agroclimatologia e o cultivo de citros. In: RODRIGUES, O.; VIÉGAS, F.; POMPEU JUNIOR, J. & AMARO, A.A., eds. Citricultura brasileira 2.ed. Campinas, Fundaçăo Cargill, 1991. p.153-195.
- POMPEU JUNIOR, J. Porta-enxertos. In: RODRIGUES, O.; VIÉGAS, F.; POMPEU JUNIOR, & AMARO, A.A.J. eds. Citricultura brasileira, 2.ed. Campinas, Fundaçăo Cargill, 1991. p.265-280.
- POMPEU JUNIOR, J.; FIGUEIREDO, J.O.; TEÓFILO SOBRINHO, J.; JORGE, J.P.N. & SALIBE, A.A. Comportamento da laranjeira 'Valęncia' em 18 porta-enxertos. In: CONGRESSO BRASILEIRO DE FRUTICULTURA, 6., 1981. Anais Recife, Sociedade Brasileira de Fruticultura, 1981. v.2, p.530-541.
- RAMOS, J.W. & KAUFMANN, M.R. Hydraulic resistance of rough lemon roots. Physiologia Plantarum, Copenhagen, 45:311-314, 1976.
- REUTHER, W. Citrus. In: ALVIM, P.T. & KOZLOWSKI, T. T. Ecophysiology of tropical crops New York, Academic Press, 1977. p. 409-440.
- SINCLAIR, T.R. & ALlEN, J.r. Carbon dioxide and water vapour exchange of leaves on field-grown citrus trees. Journal of Experimental Botany, Oxford, 33:1166-1175, 1982.
- STEVENS JR., G.A. & WESTWOOD, M.N. Fruit set and cytokinin-like activity in the xylem sap of sweet cherry as affected by rootstocks. Physiologia Plantarum, Copenhagen, 61:464-468, 1984.
- SYVERTSEN, J.P. Light acclimation in citrus leaves. II. CO2 assimilation and light, water, and nitrogen use efficiency. Journal of American Society of Horticultural Science, New York, 109:812-817, 1984.
- SYVERTSEN, J.P. & GRAHAM, J.H. Hydraulic conductivity of roots, mineral nutrition, and leaf gas exchange of Citrus rootstocks. Physiologia Plantarum, Copenhagen, 61: 464-468, 1985.
- TENNANT, D. A test of a line intersect method of estimating root length. Journal of Ecology, London, 63: 995-1001, 1975.
- TEÓFILO SOBRINHO, J. Variedades copas e porta-enxertos para os citros. In: MENTEN, J.O.M., DOURADO NETO, D. & TORRADO, P.V. Curso Intensivo de Citricultura Piracicaba, ESALQ, 1991. p.25-36.
- TURNER, N.C.; SCHULZE, E. D. & GOLLAN, T. The response of stomata and leaf gas exchange to vapor pressure deficits and soil water content. II. In the mesophytic herbaceous species Heliantus annuus Oecologia, Berlin, 65:348-355, 1985.
- VU, J.C.V.; YELENOSKY, G. & BAUSHER, M.G. CO2 exchange rate, stomatal conductance, and transpiration in attached leaves of Valencia orange. HortScience, Alexandria, 21:143-144, 1986.
- WARDOWSKI, W.F.; NAGY, S. & GRIERSON, W. Fresh Citrus Fruits. 1.ed. An AVI Book, New York, 1986. 571 p.
- WUTSCHER, H.K. Citrus rootstocks. Horticultural Reviews, Westport, 1:237-269, 1979.
- WUTSCHER, H.K. & BISTLINE, F. W. Performance of `Hamlin' orange on 30 citrus rootstocks in southern Florida. Journal of American Society of Horticultural Science, 113:493-497, 1988.

Datas de Publicação
-
Publicação nesta coleção
16 Jun 1999 -
Data do Fascículo
1998
Histórico
-
Recebido
02 Jul 1997 -
Aceito
31 Jan 1998