Abstracts
Eurhinus magnificus Gyllenhal has been collected in south Florida, presumably introduced through trade with countries in its native range. Very little information has been published on the biology or taxonomy of this insect. We conducted studies to investigate various aspects of its life history and host plant associations. The pre-imaginal life stages of E. magnificus are described for the first time. Dimensions of the adult, egg, larval, and pupal stages are also provided; head capsule measurement revealed five larval instars. All life stages of E. magnificus were collected at several sites in Broward and Miami-Dade counties from the host plant Cissus verticillata (L.) Nicolson & Jarvis (Vitaceae). Eggs were laid singly within the succulent, young subterminal portion of the host plant stem, one or two occurring between two nodes. Gall formation was apparent by the first to third instar and continued to increase in size until pupation which occurred within the gall. Predators and pathogens appeared to be responsible for considerable mortality in the field; there was no evidence of parasitism of any of the life stages. Greenhouse studies were undertaken to determine E. magnificus development time and host specificity. Adult weevils attacked grape cultivars (Vitis spp.) and feeding opened the stems to fungal agents but no physical evidence of larval development was apparent on grapes.
Baridinae; Cissus; development; Florida
Se ha recolectado, en el sur de Florida, a Eurhinus magnificus Gyllenhal. Se efectuaron estudios de investigación sobre varios aspectos del ciclo biológico, y asociaciones entre el picudo y plantas hospederas. Se describen aquí por primera vez los estados inmaduros de E. magnificus. Se dan a conocer las dimensiones de los estados de adulto, huevo, larva y pupa. Las medidas de la cápsula cefálica revelaron que hay cinco estadios en esta especie. Todos los estados de E. magnificus fueron colectados en la planta hospedera Cissus verticillata (L.) Nicholson y C.E. Jarvis (Vitaceae) en varios lugares localizados en los condados de Broward y Miami-Dade. Los picudos depositan un o dos huevos dentro de la parte más tierna del tallo que se encuentra entre dos yemas laterales localizadas en la región subterminal de la planta hospedera. En esta región, aparece una agalla, la cual es más visible cuando la larva esta entre el primer y tercer estadios. La agalla crece en tamaño hasta empupamiento, el cual ocurre dentro de esta. Los depredadores y patógenos parecen ser responsables por una gran mortalidad de estados inmaduros en el campo; En el campo, no se observó ningun indice de parasitismo en esta especie. Se llevaron a cabo estudios en casa de malla con el fin de determinar el tiempo de desarrollo y especificidad de hospederos de E. magnificus. Los picudos atacaron cultivares de uva (Vitis spp. - Vitaceae). No se observó el desarrollo larvario de E. magnificus en Vitis L.
Baridinae; Cissus; picudo; desarrollo; Florida
SYSTEMATICS, MORPHOLOGY AND PHYSIOLOGY
Life history and larval morphology of Eurhinus magnificus Gyllenhal (Coleoptera: Curculionidae), a new weevil to the United States
Ciclo biológico y morfología larval de Eurhinus magnificus Gyllenhal (Coleoptera: Curculionidae), un nuevo picudo de los Estados Unidos
Bryan J. UlmerI; Rita E. DuncanI; Jens PrenaII; Jorge E. PeñaI
IDept. Entomology and Nematology, Tropical Research and Education Center, Univ. Florida, 18905 SW 280th Street Homestead, Fl 33031
IIMozartstr. 24, 18069 Rostock, Germany
ABSTRACT
Eurhinus magnificus Gyllenhal has been collected in south Florida, presumably introduced through trade with countries in its native range. Very little information has been published on the biology or taxonomy of this insect. We conducted studies to investigate various aspects of its life history and host plant associations. The pre-imaginal life stages of E. magnificus are described for the first time. Dimensions of the adult, egg, larval, and pupal stages are also provided; head capsule measurement revealed five larval instars. All life stages of E. magnificus were collected at several sites in Broward and Miami-Dade counties from the host plant Cissus verticillata (L.) Nicolson & Jarvis (Vitaceae). Eggs were laid singly within the succulent, young subterminal portion of the host plant stem, one or two occurring between two nodes. Gall formation was apparent by the first to third instar and continued to increase in size until pupation which occurred within the gall. Predators and pathogens appeared to be responsible for considerable mortality in the field; there was no evidence of parasitism of any of the life stages. Greenhouse studies were undertaken to determine E. magnificus development time and host specificity. Adult weevils attacked grape cultivars (Vitis spp.) and feeding opened the stems to fungal agents but no physical evidence of larval development was apparent on grapes.
Key words: Baridinae, Cissus, development, Florida
RESUMEN
Se ha recolectado, en el sur de Florida, a Eurhinus magnificus Gyllenhal. Se efectuaron estudios de investigación sobre varios aspectos del ciclo biológico, y asociaciones entre el picudo y plantas hospederas. Se describen aquí por primera vez los estados inmaduros de E. magnificus. Se dan a conocer las dimensiones de los estados de adulto, huevo, larva y pupa. Las medidas de la cápsula cefálica revelaron que hay cinco estadios en esta especie. Todos los estados de E. magnificus fueron colectados en la planta hospedera Cissus verticillata (L.) Nicholson y C.E. Jarvis (Vitaceae) en varios lugares localizados en los condados de Broward y Miami-Dade. Los picudos depositan un o dos huevos dentro de la parte más tierna del tallo que se encuentra entre dos yemas laterales localizadas en la región subterminal de la planta hospedera. En esta región, aparece una agalla, la cual es más visible cuando la larva esta entre el primer y tercer estadios. La agalla crece en tamaño hasta empupamiento, el cual ocurre dentro de esta. Los depredadores y patógenos parecen ser responsables por una gran mortalidad de estados inmaduros en el campo; En el campo, no se observó ningun indice de parasitismo en esta especie. Se llevaron a cabo estudios en casa de malla con el fin de determinar el tiempo de desarrollo y especificidad de hospederos de E. magnificus. Los picudos atacaron cultivares de uva (Vitis spp. - Vitaceae). No se observó el desarrollo larvario de E. magnificus en Vitis L.
Palabras-clave: Baridinae, Cissus, picudo, desarrollo, Florida
Eurhinus magnificus Gyllenhal was first reported in Florida in 2002 when a single specimen was found in an ornamental nursery in Broward County (Feiber 2002). It was collected again in 2003 near Homestead, Miami-Dade County, and was also intercepted in a shipment of bananas from Costa Rica (Fowler 2004). It is likely that this weevil has been inadvertently imported into Florida through trade in live plants or plant products. During the present study in 2005, all life stages of E. magnificus were collected repeatedly in both Broward and Miami-Dade Counties.
E. magnificus is a member of the Curculionidae-Baridinae. The original spelling of the genus, Eurhin Illiger 1807, was suppressed by the plenary power of the International Commission on Zoological Nomenclature (Melville 1985), thereby following the arguments of Zimmerman and Thompson (1983) for resolving a nomenclatural conflict between two homonymously used family-group names. The genus includes 23 species that are widely distributed in Central and South America (Vaurie 1982). Eurhinus magnificus occurs in Belize, Costa Rica, Guatemala, Honduras, Mexico, Nicaragua and Panama (Vaurie 1982, Schall 2002). It is a robust weevil similar in body form to other Eurhinus weevils (Bondar 1948) and like many of the other species, E. magnificus adults are brilliantly colored. The entire body is a vibrant metallic blue-green with areas of metallic red-copper on the humeri and apex of the elytra and on the pronotum, rostrum, and legs. Little is known about adult behavior, however, it is known that at least two species deposit eggs in the stem of host plants and that developing larvae feed on plant juices within a gall formed by the developing larva (Bondar 1948).
Like other members of the genus, E. magnificus larvae induce galls on the stems of Cissus spp. (Vitaceae) (Bondar 1948). Vernonia, Andira and Mikania spp. have also been suggested as host plants (Vaurie 1982), however, the only verified host plant of E. magnificus in Florida is Cissus verticillata (L.) Nicolson & Jarvis (Vitaceae). Florida contains native species of all four genera which are also represented by species occurring over a wide range in the United States (Fowler 2004, USDA 2006). C. verticillata is known by the common names of possum grape vine, princess vine, and season vine, and is widely distributed in the Caribbean, Central and South America, and Mexico (Lombardi 2000); it is considered native to Florida in the United States (USDA 2005). Though C. verticillata is sometimes planted as an ornamental, it is a prolific perennial vine that is generally considered a weedy species when occurring with various ornamental and fruit crops in south Florida (Futch & Hall 2003).
The aim of the present study was to investigate the occurrence of E. magnificus in south Florida and gain insight into its life history. Very little is known about the biology of this exotic weevil and nothing is known of its abundance or potential impact in Florida. Field collections of both adult and immature stages were made in Broward and Miami-Dade counties. Various aspects of host plant interaction were considered including adult feeding, oviposition, and the relationship between larval development and host plant phenology. The growth of E. magnificus was studied to determine the number of larval instars and development time. We also describe the pre-imaginal life stages of E. magnificus for the first time. Host plant specificity and the vulnerability of commercial grape cultivars (Vitis spp.) to E. magnificus were also considered.
Materials and Methods
Field collection and processing. The occurrence of E. magnificus in south Florida was investigated from Homestead, Miami-Dade County (25º 29'45 N, 80º 23'42 W) to Fort Lauderdale, Broward County (26º 04'38 N, 80º 18'38 W). Various habitats including ornamental nurseries, citrus groves, roadside ditches, fence lines, and manicured gardens were searched for the presence of adult weevils and C. verticillata galls containing immature stages of E. magnificus. On 30 June 2005 and approximately weekly thereafter until early November 2005, adult specimens and host plant material were collected in the field. Adults were collected individually by hand directly from host plants or occasionally by beating host plants over an umbrella which caught adults as they fell from the foliage. Immature stages were located by visually inspecting host plant stems in the field. When the presence of immature stages was indicated by an oviposition cavity or a gall on the host plant stem, that portion of the plant was cut to include at least one node on either side of the injury and returned to the laboratory. The stems were dissected under a dissecting microscope and examined for the presence of E. magnificus pre-imaginal life stages.
Measurements were made of E. magnificus adults, eggs, head capsule widths of the different larval instars, and body widths of larvae using an ocular micrometer mounted in a dissecting microscope. Adult (n = 15), pupal (n = 9) and egg (n = 5) measurements were made on their longest and widest axes. Larval (n = 74) head capsules were measured dorsally along the widest axis. The crescent shape of the larvae did not lend itself to length measurements without damaging the larvae, thus larval width was measured at the widest point on the body for each specimen.
Pre-imaginal morphology. Larvae recovered from host plants were boiled for 1 min and preserved in 75% ethanol. The morphology of the immature stages was studied as described by May (1994), and her terminology (p. 723-726) is followed here. The numbering of the setae on the head and mouth parts proceeds from base to front.
Development and host plant interaction. To investigate host plant interactions, C. verticillata was collected in the field and grown in an outdoor greenhouse. Cuttings of approximately 30-45 cm were placed in water until roots formed. When roots were well established (15-30 cm long) the plants were transplanted into potting soil in 15-cm diameter plastic pots and maintained in the greenhouse. Host plants were exposed to mated pairs of field collected adults for periods of 37 d in 70 x 70 x 70 cm mesh cages in the greenhouse after which all adult weevils were removed from the host plants. Feeding and oviposition habits of adult weevils were observed during exposure. After exposure, host plants were maintained in the greenhouse and galls containing immature stages of E. magnificus were periodically dissected to determine developmental times for the immature stages. The greenhouse experiment was conducted from mid-October, 2005 to January 20, 2006. The mean air temperature in the greenhouse was 20.0ºC; the maximum temperature was 38.6ºC and the minimum temperature was 2.4ºC. Air temperature was recorded using a temperature probe (StowAway®TidbitT® made by Onset) hanging 4 cm below the top of the cage. Adult longevity was determined by placing newly emerged field collected adults in 30 x 30 x 30 cm Plexiglass cages in the laboratory. The weevils were provided water via a cotton wick and a food source consisting of cuttings of C. verticillata, including several leaves and 30-45 cm of stem material, and held in a room maintained at 24ºC and approximately 70% RH. Adults were checked daily for signs of life and the time of death was recorded.
To assess weevil development in relation to host plant phenology, infested stems of C. verticillata were collected from the field (as above). Each stem containing immature stages of E. magnificus was measured with a caliper (to the nearest 0.1 mm) adjacent to the node at the proximal end of the infested internode. The diameter of the gall at the widest point was also measured and the stem dissected to determine the developmental stage of the weevil. Larvae were examined for signs of life and parasitism. Stem and gall diameter means for each life stage were compared by Tukey's Honestly Significant Differences (HSD) test following a significant ANOVA, a Linear Regression was also applied to evaluate the relationship between host plant stem and gall diameter and the development stage of E. magnificus (Statistix® 8 Analytical Software 2003).
Host plant specificity. To determine host plant specificity, field collected adults were offered commercial grape cultivars including Carlos, Noble, Welder (Vitis rotundifolia Michx.) and L.K. Emerald (V. vinifera L.). Grapes were planted singly in 15-cm diameter pots (as above). The plants were 1-2 m from the soil surface to the tip of the vine at transplanting in August 2005. Two plants of each cultivar were offered individually to a mated pair of E. magnificus adults in the greenhouse for 14 d in mesh cages (as above) after which the weevils were removed. After exposure, damage and gall formation was documented weekly and the grape plants were held in the greenhouse for 67 d to 95 d before being dissected under a microscope to determine the success of E. magnificus on each cultivar.
Voucher specimens. Specimens from each collecting period were sent to FDACS-DPI and identified by M. C. Thomas. Vouchers are deposited in the Florida State Collection of Arthropods.
Results
Field collections. June to November 2005, all life stages of E. magnificus were collected at several sites in Broward and Miami-Dade counties from the host plant C. verticillata. Adults and galls were found in various habitats where C. verticillata occurred including fence lines, ditches, citrus groves and ornamental nurseries. The number of specimens collected by two individuals in 1h ranged from zero to 74 adult weevils and over 100 galls depending on the location and environmental conditions. A single adult was also found on pigmy palm, Phoenix robelenii O'Brien (Arecaceae), and one was collected from avocado, Persea americana Mill. (Lauraceae) foliage. However, no evidence of feeding or oviposition were observed on these two plants and it is likely that the collections were incidental and they are not true host plants; C. verticillata is a prolific vine that is often closely associated and intermingled with other plants in the groves and nurseries in south Florida. Host plant associations reported for E. magnificus and several other members of the genus are included in Table 1.
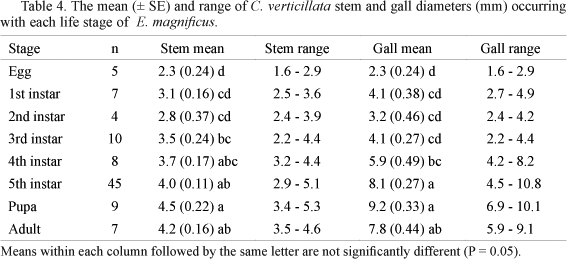
Mean length and width of field collected E. magnificus adult, egg, and pupal stages are listed in Table 2. Head capsule measurement revealed that E. magnificus completes five larval instars before pupation (Table 3).
Pre-imaginal morphology. Description of larva: Habitus: maximum size 9.0/mm x 3.8/mm, body stout, curved, greatest width near middle, tapered to both ends (Fig. 1A); segmental lobes protruding, abdominal segments IVII with three tergal folds, prodorsal folds 2-lobed, postdorsal folds 6-lobed, caudal segments unmodified; cuticle smooth, finely punctate, setae brown. Head: subprognath, amber-brown with v-shaped pattern, frontal margin and endocarina dark; head capsule as long as wide, greatest width behind middle, gradually narrowed to front; postoccipital condyle triangular; epicranial ridge absent, frontal suture slightly sinuate, endocarina 0.7x length of frons, two pairs of stemmata; frontal setae 1, 2, 4 and 5 long, 3 short; dorsal epicranial setae 15 long; 4 or less posterior epicranial sensilli each side (Fig. 1B); lateral epicranial setae 1 and 2 long; ventral epicranial seta 1 long, 2 short (Fig. 1C); accessory appendage of basal antennal segment conical, 2x longer than wide, 56 minute basal sensilli (Fig. 1D); clypeus 3.5x wider than long, subconical, clypeal setae 1 and 2 moderately long; labrum with blunt anteromedian lobe, three pairs of variously long labral setae (Fig. 1E), tormae separate, bar-shaped, curved, epipharyngeal lining with two pairs of short anteromedian setae and one pair of anteromedian sensilli, three moderately long anterolateral setae, two pairs of short median setae and one sensillus in between (Fig. 1F); maxilla with palpus 2-segmented, basal segment with one short seta and two sensilli, mala with five distal setae and comb of 67 dorsal setae; labial palpus 2-segmented, premental sclerite basally produced, one pair of long prelabial setae, postlabial seta 3 shorter than setae 1 and 2, ligula with two pairs of short setae and two pairs of sensilli (Fig. 1G); mandible with two apical teeth, setae arranged longitudinally (Fig. 1H). Thorax: creamy white without pigments; 10 pairs of variously long pronotal setae, two long pleural setae, two long and four shorter pedal setae, one pair of short sternal setae; thoracic spiracle subcircular, approximately 1.5x larger than abdominal spiracles, two annulated air-tubes; meso- and metathorax with one pair of long prodorsal and four pairs of variously long postdorsal setae, one moderately long dorsolateral seta, one long and one short alar seta, one long epipleural seta, one long pleural seta, two long and 4(5) more-or-less short pedal setae, one pair of short sternal setae. Abdomen: creamy white without pigments; spiracles of segments IVIII subcircular, with two annulated air-tubes; segments IVII with five pairs of postdorsal setae of unequal length, VIII with three pairs; IVIII with one pair of short prodorsal setae, two short spiracular setae, two epipleural setae of unequal length, two pleural setae of unequal length, one short laterosternal seta, two pairs of short sternal setae; segment IX with two pairs of dorsal setae, two pairs of ventral setae and one lateral seta; anus terminal, 4-lobed, lateral lobe with two short setae. Alimentary canal: cardiac valve with ring of mycetomes, ventriculus with approximately 20 short vermiform gastric caeca each side, Malpighian tubules grouped 4+2 at origin, cryptonephridium well-developed, rectal bracon ring-shaped. Material: 10 final instar (5 dissected), one first instar and two third instar larvae collected from galls of C. verticillata on 29 September 2005 at Fort Lauderdale, FL.
Description of pupa: Habitus: 8.1 mm x 3.8 mm, stout (Fig. 1I); setae translucent. Head: retracted, barely visible in dorsal view, one pair of setae on vertex, one pair of setae above eyes; rostrum reaching slightly beyond fore coxae, one pair of setae between eyes, one pair of setae in distal one-third of rostrum. Thorax: pronotum transverse, greatest width at base, sides rounded toward front, apex subtubulate, basal margin very slightly produced, disk without depression, five pairs of dorsal and four pairs of lateral setae (Fig. 1K); mesonotum with three pairs of setae, scutellum distinct, transverse, slightly elevated; metanotum not sulcate medially, two pairs of setae; pterotheca I smooth, finely punctate, tubercles absent, pterotheca II as long as pterotheca I; femora with one seta each. Abdomen: all setae set on tubercles; segments IVII with two pairs of postdorsal and two pleural setae, segment VIII with two pairs of postdorsal setae, one pleural seta and one pair of ventral setae, segment IX with one pair of caudally oriented pseudocerci and one pair of short setae. Material: two specimens with same collecting data as larvae described above.
Development and host plant interaction. Gall dissections at various intervals showed that 20-25 d after oviposition E. magnificus larvae had reached the second instar (n = 3) and by 40-45 d after oviposition larvae had reached the fifth instar (n = 2) under greenhouse conditions. Pupation occurred after 61-67 d (n = 1) and adult emergence was 80-86 d (n = 1) after oviposition. The experiment was conducted in an outdoor greenhouse beginning October 26, 2005.
Adults emerging from field collected galls, provided host plant material and water, survived 19-47 d (mean 32.0 ± 7.0 d, n = 4) from the time of emergence under laboratory conditions.
The growth stages (diameter) of the host plant stem in which the different life stages of E. magnificus were found were significantly different (F7,87 = 9.2, P < 0.001, r2 = 0.38) (Table 4). Eggs to second instar larvae were found in relatively young parts of the stem, small in diameter, while late instar larvae, pupae and adults were found in significantly larger stems. The size of the gall occupied by different life stages was also significantly different (F7,87 = 28.1, P < 0.001, r2 = 0.58). Gall formation was not detectable at the egg stage, but became apparent at the first to third instar. By the fourth instar the gall was substantially larger than the host plant stem. Gall size was maximal at the pupal stage (9.2 ± 0.3 mm), decreasing slightly by adult emergence (Table 4).
In 44% of the infested stems collected in the field E. magnificus did not survive. The remains of all life stages from egg to adult were found within host plant stems, however, the mortality factors for each life stage were not clear. The galleries created by deceased E. magnificus larvae were frequently infested with various mites and ants. Many of the cadavers were also enveloped with fungal agents. In many instances as the gall increased in size and dried, the protective plant tissue cracked and split, possibly leaving immature weevils more vulnerable to these environmental hazards. There was no evidence of parasitism of any of the life stages. Forty-one of the 95 living E. magnificus specimens collected from galls occurred singly between two nodes in the host plant stem. The remaining specimens were found in pairs 0.5 cm to 2.5 cm apart. When two specimens occurred within the same internode they were generally indistinguishable in size and developmental stage, presumably oviposited by the same female at approximately the same time.
Host plant specificity. After 14 d exposure to E. magnificus, grape cultivars were examined for signs of damage. Rasping on the stems characteristic of the injury observed for E. magnificus feeding and oviposition on C. verticillata was observed on the grape cultivars Carlos, Noble, and Welder. There was no indication of injury on cv. L.K. Emerald. Fifteen days after exposure, galls were observed on the Carlos, Welder, and Noble cultivars. Given the small number of galls, the stems were not dissected in anticipation of larval development. Sixty-seven days after exposure, galls were still apparent but had not developed noticeably beyond the point they were at on day 15. The galls measured 3.1-3.3 mm on cv. Carlos (n = 3), 1.9-2.7 mm on cv. Welder (n = 4), and 3.3 mm on cv. Noble (n = 1), approximately 1.5-2x the diameter of the stem they were located on for each cultivar. Galls on the three cultivars were dissected under a microscope. No larvae were found alive. If oviposition occurred and larvae hatched, it seems they died early in development. The galls were infested with fungal agents and mites and there were ants on the plants. It is possible that young larvae were consumed or removed. The stems and leaf petioles of the grape cultivars tested were relatively woody and rigid compared to those of similar size on the host plant C. verticillata which were more supple and succulent.
Discussion
E. magnificus was abundant and successfully reproducing in various habitats where the host plant C. verticillata was present across Miami-Dade and Broward Counties. After first being reported in Florida in 2002 (Feiber 2002), it appears to be established and flourishing in the southeast region of the state. C. verticillata supports the entire life cycle of E. magnificus. Eggs are laid within the stem where larvae hatch and begin to feed. The larvae complete five instars within a gall formed at the location of oviposition before pupating. Adults emerge from the host plant gall to feed on C. verticillata, mate and oviposit. There was no evidence of larval or adult feeding or oviposition on any other plants occurring in the company of E. magnificus.
Adults were observed feeding on the outer layers of host plant stems and also within cavities created in the stems and the leaf petioles. Some feeding also occurred on leaf blades but this was restricted to that portion of the leaf directly around the attachment point of the petiole. Oviposition generally occurred within actively growing portions of the stem (< 3 mm in diameter) that were green and succulent. Females created a cavity into the pith of the stem with their rostrum and oviposited a single egg into the cavity. One or two eggs were laid within a single host plant internode with similar frequency, more than two eggs in a single internode were never observed in the field or the greenhouse. By the second instar, gall formation was usually evident and progressed to become approximately twice the diameter of the stem by pupation. Galls generally began to dry out and marginally reduce in size before adult emergence.
A single specimen of E. magnificus required approximately 83 days to develop from egg to adult, however, our study was conducted during the coldest period of the year. It is likely that the development time of E. magnificus would be greatly reduced during the summer months when daily mean minimum and mean maximum temperatures at Homestead, FL are 7.0ºC and 8.7ºC higher, respectively, than they were during the months our experiments were conducted (Southeast Regional Climate Center 2005). Our data may not reflect development under optimal temperatures for development, however, the presence of adults in the field and larval development observed during winter indicate that E. magnificus is capable of reproducing throughout the year in south Florida.
The impact of E. magnificus on C. verticillata was not directly measured. In general the host plants appeared healthy and actively growing beyond the point of gall formation. However, the development of E. magnificus within the stem caused a significant malformation and often had an obvious negative impact on the host plant; occasionally the gall and resulting damage coupled with environmental conditions resulted in severing the stem. Negative effects on host plants were most apparent when infestations were high, often resulting in numerous galls on individual plants. Given the weed status of C. verticillata among ornamental and fruit crops in south Florida (Futch & Hall 2003), E. magnificus could be a welcome biological control agent for this plant. However, populations would need to increase above those observed in the present study before E. magnificus had an impact on the field populations of C. verticillata.
Stem dissections indicated a high mortality rate within the galls of host plants, successful development of E. magnificus in the field appeared to be significantly reduced by predation and fungal agents. However, the examination of approximately 200 specimens of E. magnificus in the present study revealed no evidence of parasitism for any life stage. There are no records of parasitism for E. magnificus in its native range. Capitonius tricolorvalvus Ent (Hymenoptera: Braconidae) was reared from stem galls of C. verticillata in Costa Rica, the host was not identified but presumed to be a beetle (Ent & Shaw 1999). Further sampling is needed to establish the parasitoid fauna, or lack of, associated with this weevil in Florida and its native range.
Eurhinus magnificus is not known to be a pest of commercial grapes and is not considered a threat to attack Vitis spp. (Fowler 2004). However, our preliminary experiments showed that E. magnificus adults did accept grape cultivars as a feeding substrate. The limited feeding damage and lack of evidence for oviposition or larval development indicate that the grape cultivars we tested were not suitable hosts for E. magnificus. However, three of the four cultivars tested were attacked by adult weevils in a no-choice situation and it is conceivable that this weevil could develop on other grape cultivars under different conditions. Additional study investigating a wide range of grape cultivars varying in physical and chemical characteristics should be undertaken before eliminating Vitis spp. as suitable host plants. Though E. magnificus did not develop within grape plants, the damage and resulting galls did open the plants to fungal agents that spread into the pith from the point of infection.
E. magnificus is established in southeast Florida. This attractive exotic weevil is flourishing on the host plant C. verticillata throughout Miami-Dade and Broward Counties. The impact of the weevil on this native plant species and on other potential hosts, such as Vernonia spp. and Vitis spp., is not discernible at this early stage of colonization but necessitates future research. Further work is also required to better understand the population biology of E. magnificus and to establish its potential for range expansion in the United States.
Acknowledgments
We are grateful to M. C. Thomas for specimen identification and critical review of the manuscript. We also thank D. Long, J. Alegría, and Z. Alegría for technical assistance. Special thanks to Henry Hespenheide, Anne Howden, Kenji Nishida, Charles W. O'Brien, Frode Oedegaard, Wendy Porras and Henry Stockwell for providing unpublished observations on plant associations and Barry Hammel for identification of Cupania guatemalensis.
Received 10/IV/06. Accepted 11/XI/06.
- Bondar, G. 1948. Notas entomologicas da Baía XX. Rev. Entomol., Rio de J. 19(1-2): 1-54.
- Ent, L.J. van der & S.R. Shaw. 1999. A new species of Capitonius (Hymenoptera: Braconidae) from Costa Rica with rearing records. Pan-Pacific Entomol. 75: 112-120.
- Feiber, D. 2002. Plant industry update. Florida Department of Agriculture and Consumer Services. Fall 2002, p.4. Download at: www.doacs.state.fl.us/pi/images/F02newsletter.pdf
- Fowler, L. 2004. Eurhin magnificus Gyllenhal: Weevil Coleoptera/Curculionidae. New Pest Advisory Group (NPAG). Plant Epidemiology Risk Analysis Laboratory, Center for Plant Health Science and Technology. Report 040305, p.1-2.
- Futch, S.H. & D.W. Hall. 2003. Identification of vine weeds in Florida citrus. Horticultural Sciences Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida. Document HS926, August 2003. Download at http://edis.ifas.ufl.edu
- Lombardi, J.A. 2000. Vitaceae: Generos Ampelocissus, Ampelopsis, e Cissus Flora Neotr. Monograph. 80: 200.
- Maes, J.-M. 1999. Insectos de Nicaragua, p.486-1169. V. II. Setab BOSAWAS, MARENA, Managua, Nicaragua, 1898p.
- May, B.M. 1994. An introduction to the immature stages of Australian Curculionoidea, p. 365-728. In E.C. Zimmerman, Australian weevils, v. II, VIII, 755p.
- Melville, R.V. 1985. Opinion 1352 Eurhinus Schönherr, 1825 (Insecta, Coleoptera): ruled as a justified emendation of Eurhin Illiger, 1807. Bull. Zool. Nom. 42: 287-290.
- Schall, R. 2002. Eurhin magnificus A Central American weevil. New Pest Advisory Group (NPAG) Data. p.9. In Fowler, L. 2004. Eurhin magnificus Gyllenhal: Weevil Coleoptera/Curculionidae. New Pest Advisory Group (NPAG). Plant Epidemiology Risk Analysis Laboratory, Center for Plant Health Science and Technology. Report 040305.
- Silva, A., G. d'Araújo e, C. Rory Gonçalves, D. Monteiro Galvão, A.J. Lobo Gonçalves, J. Gomes, M. do Nascimento Silva, & L. de Simoni. 1968. Quarto catálogo dos insetos que vivem nas plantas do Brasil. Seus parasitos e predatores, v. 1 (2). Ministério da Agricultura, Rio de J., III-XXVI, 622p.
- Southeast Regional Climate Center. 2005. Historical climate summaries for Florida. Download at: www.dnr.state.sc.us/climate/sercc/climateinfo/historical/historical_fl.html (October 13, 2005).
- Statistix® 8 Analytical Software. 2003. Statistix® 8 User's Manual. Tallahassee Florida: Analytical Software. ISBN 1-881789-06-3.
- United States Department of Agriculture. 2005. Cissus verticillata (L.) Nicolson & C.E. Jarvis. Germplasm Resources Information Network (GRIN) Taxonomy for plants. Download at: http://www.ars-grin.gov/~sbmljw/cgi-bin/taxon.pl?310369 (December 8, 2005). Place of publication: Taxon: 46: 430. 1997.
- United States Department of Agriculture. 2006. Natural Resources Conservation Service. Plants Database. Download at: http://plants.usda.gov/ (March 31, 2006)
- Vaurie, P. 1982. Revision of Neotropical Eurhin (Coleoptera, Curculionidae, Baridinae). Novitates 2753: 1-44.
- Zimmerman, E.C. & R.T. Thompson. 1983. On family group names based upon Eurhin, Eurhinus and Eurhynchus (Coleoptera). Bull. Zool. Nom. 40: 45-52.
Publication Dates
-
Publication in this collection
09 Aug 2007 -
Date of issue
June 2007
History
-
Received
10 Apr 2006 -
Accepted
11 Nov 2006