ABSTRACT
Avian leukosis virus subgroup J (ALV-J), a member of the retroviridae family, can infect both broilers and layers and induce a spectrum of different neoplasms, resulting in serious economic losses in poultry production. Epigallocatechin-3-gallate (EGCG), the major constituent of green tea, has demonstrated remarkable anti-inflammatory and cancer chemopreventive effects in many animal tumor bioassays, cell culture systems and epidemiological studies. To assess the antiviral effects of EGCG on ALV-J-induced cell apoptosis in vitro, DF-1 cells were treated with different EGCG concentrations (0, 5, 10, 20 and 40 µg/mL), and their antiviral effects were examined at different time points (0, 24, 48, 72 and 96 h) using a variety of assays. EGCG alleviated the ALV-J-induced apoptosis in a dose-dependent manner. Because high concentrations (20 and 40 µg/mL) inhibited DF-1 cell growth, and low concentration (5 µg/mL) did not suppress the ALV-J virus, 10 µg/mL was the most appropriate concentration. After 96 h of incubation, 10 µg/mL EGCG improved the ALV-J-triggered suppression of the nuclear transcription factor system by enhancing cytoplasmic NF-κB p50/p65 expression and inhibiting nuclear NF-κB p50/p65 expression, resulting in decreased cell apoptosis. These results demonstrated that EGCG inhibited ALV-J-induced apoptosis in DF-1 cells in a dose-dependent manner via the NF-κB signaling pathway, and that 10 µg/mL EGCG is the optimal concentration, which may be useful for therapeutic drug design.
Keywords: ALV-J; EGCG; NF-kB; DF-1 cell; apoptosis
INTRODUCTION
Avian leukosis virus (ALV), which belongs to the retroviridae family, induces benign and malignant neoplasms in chickens (Silva et al., 2007). Chicken ALVs can be divided into exogenous (subgroups A-D and J) and endogenous (subgroup E) viruses based on the viral interference patterns, virus neutralization and host range (Gao et al., 2015). ALV subgroup J (ALV-J) was first isolated from meat-type chickens by Payne et al. in the United Kingdom in 1988, and since then this disease has become widespread, causing serious economic losses in poultry production (Payne et al., 1991, Li et al., 2015b). Although ALV-J has been successfully contained or eradicated from the breeding flocks in some countries, this virus gained a strong foothold in China, inducing various tumors in commercial layer flocks and has even spread to the local chicken breeds in recent years (Fan et al., 2012). Enhancement of these diseases by concomitant ALV-J infection is most likely the consequence of immunosuppression (Gao et al. 2015). So far, an effective commercial vaccine against ALV-J is not available, and eradication of ALV-J has been difficult, because of substantial genetic and antigenic variation among ALV-J isolates, as well as high levels of vertical and horizontal transmission (Gao et al., 2010).
Epigallocatechin gallate (EGCG) is a polyphenolic compound abundant in green tea, which is one of the most common beverages consumed in the world (Van Aller et al., 2011). EGCG is known as an antioxidant compound, which has been found to suppress the inflammatory processes that lead to transformation, hyperproliferation, and initiation of carcinogenesis; its cancer-preventive effects are widely supported by results from epidemiological, cell culture, animal and clinical studies (Singh et al., 2011, Lecumberri et al., 2013). The mechanism of EGCG-mediated apoptosis in cancer cells is not completely understood. An in vitro cell culture study has suggested that EGCG potently induces apoptosis and promotes cell growth arrest by changing the expression of cell cycle regulatory proteins, activating killer caspases, and suppressing nuclear factor-kappa B (NF-κB) activation (Gupta et al., 2004). Recently, EGCG has been proven to have an anti-NF-κB transactivation activity in a number of human malignancies, such as colon cancer, breast cancer and lung cancer, and in chronic inflammation (Liu et al., 2016).
The transcription factor, NF-κB, consists of multiple members of the Rel family of proteins, including NF-κB1 (p105/p50), NF-κB2 (p100/p52), RelA (p65), RelB and c-Rel, which possess a conserved Rel homology domain that is responsible for dimerization, nuclear localization and DNA-binding (Prasad et al., 2010). The NF-κB pathway is activated through two distinct signaling pathways; in the classical pathway degradation of IκBa by IkB kinase (IKK) b activation triggers the translocation of various heterodimers, predominantly p65/p50, to the nucleus (Maruyama et al., 2016). Several studies have demonstrated that NF-κB is critical for the control of the cellular stress response and is involved in the regulation of cell cycle/growth, apoptosis, inflammation and immunity. Furthermore, it has been implicated in mediating proliferation and apoptosis in many types of cancer cells, such as human liver cancer cells, gastric cancer cells, breast cancer cells (Kim et al., 2011, Yang et al., 2012, Sun et al., 2015).
Therefore, the aim of this study was to evaluate the effect of EGCG on DF-1 cells infected with ALV-J, and to determine whether its antiviral effect is via the NF-κB signaling pathway. Our findings provide useful information for elucidating the mechanism of the EGCG effect on ALV-J-induced cell apoptosis.
MATERIALS AND METHODS
Materials
A purified EGCG preparation (> 99% pure) was extracted by the Tea Research Institute, Chongqing Academy of Agricultural Science. EGCG was dissolved in phosphate-buffered saline (PBS; pH 7.4), and used to treat DF-1 cells.
Cell culture
The DF-1 chicken embryo fibroblast cell line, free of endogenous sequences related to avian sarcoma and leucosis viruses, was purchased from Shanghai Bogoo Bio-Technique Co. Ltd, China.
DF-1 cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; GIBCO, Shanghai, China) supplemented with 10% fetal bovine serum (FBS; GIBCO, Shanghai, China), 2 mM glutamine, 100 U/mL of penicillin and 100 mg/mL streptomycin sulfate. All cells were maintained at 37 °C in a humidified atmosphere of 95% air and 5% CO2.
Determination of the cytotoxicity of EGCG to DF-1 cells
To examine the cytotoxicity of EGCG to DF-1 cells, the MTT colorimetric assay, which determines cell viability, was used.
Cells were seeded in 96-well plates at a density of 1-2×105 cells/well at 37 °C for 48 h, and then treated with EGCG solution (final concentration: 0, 5, 10, 20 or 40 µg/mL) for 24, 48, 72 and 96 h. Then, media were removed and cells were incubated with MTT (0.5 mg/ mL; Beyotime, Shanghai, China) at 37 °C for an additional 4 h. The viable cell number/dish, which is proportional to the production of formazan after solubilization with dimethyl sulfoxide, was determined by absorbance at 490 nm using a Thermo Scientific Multiskan MK3 Microplate Reader (Thermo Fisher Scientific, Waltham, MA, USA).
Determination of the preventive effect of EGCG on ALV-J-infected DF-1 cells
To evaluate the preventive effect of EGCG on ALV-J-infected DF-1 cells, DF-1 cells were plated in 96-wll plates at a density of 5´103 cells/well. When the cells reached 80-90% confluence, the culture medium was removed and the cells were pre-incubated with EGCG at different concentrations (0, 5, 10 µg/mL) at 37 °C for 2 h. Then, the cells were infected with ALV-J strain JS09GY07 at a multiplicity of infection (MOI) of 1.0. Next, medium containing 2% FBS was added to the wells, and culturing continued for 24, 48, 72 and 96 h. The cell viability was measured by the MTT assay every 24 h. During this period, the cells were briefly treated with trypsin (0.02% trypsin + 0.25% EDTA), to remove the cell surface-bound viral particles after the 3-h infection period.
Real-time PCR
Total RNA was extracted from cells treated with 10 µg/mL EGCG using Qiagen cell and tissue RNA isolation kit (Qiagen, NY, USA) according to the manufacturer’s instructions. The RNA 6000 Nano chip assay was used with a Bio analyzer 2100 (Agilent Technologies, CA, USA), to assess the RNA integrity, quality and quantity. Total RNA samples were stored at −80°C. Reverse transcription was performed using 2 µg of total RNA by PrimeScript RT Master Mix Perfect Real Time (Takara, Dalian, China) according to the manufacturer’s instructions.
Real-time PCR primers were designed by Primer Premier 5 (for env, upstream sequence: 5¢-AGAAAGACCCGGAGAAGAC-3¢, downstream sequence: 5¢-ACACGTTTCCTGGTTGTT-3¢; for b-actin, upstream sequence: 5¢-GAGAAATTGTGCGTGACATCA-3¢, downstream sequence: 5¢-CCTGAACCTCTCATTGCCA-3¢), and the mRNA abundance of each gene was determined using the CFX96-TouchTM real-time PCR detection system (Bio-Rad, Hercules, CA, USA). Real-time PCR reactions were performed in triplicate in 10 µL containing 1 µL of cDNA, 0.5 µL of reverse and forward primers (10 µM) for each gene, 3 µL of double-distilled H2O, and 5 µL of SYBR® Premix Ex TaqTM II (Bio-Rad). The gene expression was normalized to b-actin according to the 2−ΔΔCT method.
ELISA determination of ALV-J p27 antigen
The expression of ALV-J p27 antigen was determined by ELISA (IDEXX, Beijing, China) according to the manufacturer’s protocol. All determinations were performed in triplicate. The supernatant samples were harvested from cells treated with 10 µg/mL EGCG for the indicated time, and the cytoplasmic protein lysates were prepared by rapid freeze-thaw.
Western blot analysis
After 96 h of incubation, cytosolic and nuclear protein fractions were extracted from DF-1 cells treated with 10 µg/mL EGCG by the NE-PER® Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific) according to the manufacturer’s protocol. The cytoplasmic and nuclear fractions were stored at −80 °C. The Pierce bicinchoninic acid protein assay kit (Pierce, Rockford, IL, USA) was used to measure the protein concentration according to the manufacturer’s protocol.
The proteins in the gels were electrotransferred to nitrocellulose membranes (Sigma, Shanghai, China). The membranes were blocked with 5% skim milk in PBST (PBS containing 0.1% Tween 20) for 1 h at room temperature and probed for 2 h at room temperature with the appropriate primary antibodies, including chicken monoclonal antibodies against NF-κB p105/p50 (1:500 dilution; Abcam, Cambridge, MA, USA) or NF-κB p65 (1:500 dilution; Abcam) and rabbit anti-chicken b-actin polyclonal antibody (1:1000 dilution; Abcam). The blots were washed three times with PBST and incubated with the secondary antibody, goat-anti rabbit horseradish peroxidase (HRP)-conjugated IgG (1:1000 dilution; Abcam). Target proteins were detected with enhanced chemiluminescence reagent (Zomanbio, Beijing, China) using the ChemiDoc XRS system (Bio-Rad).
Assessment of NK-κB activity
The NF-κB DNA-binding activity in nuclear and cytoplasmic extracts of DF-1 cells was determined using the Trans AM™ NF-κB Family Transcription Factor Assay Kit (Active Motif, Carlsbad, CA, USA) according to the manufacturer’s protocol. About 10 µg of the cytoplasmic or the nuclear protein extracts were incubated with immobilized oligonucleotides containing the NF-κB consensus DNA-binding site (5’-GGGACTTTCC-3’) for 1 h at room temperature. After washing, 100 µL of diluted NFκB antibodies (1:1000; p50, p52, p65, c-Rel and RelB) were added for 1 h at room temperature. After three washes, 100 µL of diluted HRP-conjugated secondary antibodies (1:1000) were added to each well for 1 h at room temperature. Then, the cells were washed three times, followed by incubation with the colorimetric solution for 5 min at room temperature in the dark. Finally, the absorbance at 490 nm was determined by the Thermo Scientific Multiskan MK3 Microplate Reader (Thermo Fisher Scientific).
Statistical analysis
Data are presented as least squares means ± SEM. All statistical analyses were performed using SPSS 17.0 (SPSS Inc., Chicago, IL, USA). One-way analysis of variance and multiple comparisons (Tukey’s test) were used to evaluate the statistical significance of differences, and values were considered statistically significant at p< 0.05.
RESULTS
The cytotoxicity of EGCG to DF-1 cells
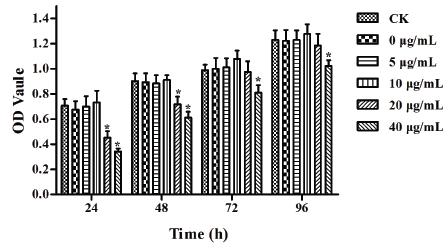
To exclude the effect of EGCG itself on DF-1 cell viability, we first examined the cytotoxicity at 24, 48, 72 and 96 h of incubation of DF-1 cells with different concentrations of EGCG (0, 5, 10, 20 and 40 µg/mL) using the MTT assay (Fig. 1). The results showed that 40 µg/mL EGCG obviously decreased the cell viability at all time points (p<0.05). Cells treated with 20 µg/mL EGCG also exhibited significantly lower viability than the control group at 24 and 48 h (p<0.05). Therefore, 5 and 10 µg/mL EGCG, which had no cytotoxic effect on DF-1 cells, were used for the following experiments.
Effect of EGCG cytotoxicity on DF-1 cell growth. The error bars represent the SEM. Differences were considered highly significant when p<0.05 (*). The data are representative of the results of three independent experiments.
The effect of EGCG on cells infected with ALV-J
To evaluate the effect of EGCG on cells infected with ALV-J, the cell viability was determined by the MTT assay (Fig. 2). The results showed that EGCG affected the cells in a dose-dependent manner, as ALV-J-infected DF-1 cells treated with 0 or 5 µg/mL EGCG exhibited remarkably lower OD values than the control (CK, blank treatment) group at all time points (p<0.05), and the OD values of the 5 µg/mL treatment group were higher than those of the 0 µg/mL treatment group (p<0.05). However, when the EGCG concentration was increased to 10 µg/mL, the OD values were similar to the CK group at all time points (p>0.05). Therefore, we used 10 µg/mL EGCG in the subsequent experiments.
Effects of genistein on ALV-J infection of DF-1 cells. The error bars represent the SEM. Differences were considered highly significant when p<0.05 (*). The data are representative of the results of three independent experiments.
Next, we validated the protective effect of 10 µg/mL EGCG on ALV-J-infected cells. To this end, we used the ALV Antigen Test Kit (ALV Ag) to examine the p27 antigen level after 96 h of incubation (Table 1). The p27 antigen was not detected in uninfected DF-1 cells at all time points. As expected, the cells infected with ALV-J exhibited a high level of p27 antigen at all time points. In the ALV-J-infected cells treated with 10 µg/mL EGCG the level of p27 antigen was significantly lower than that in ALV-J-infected cells without EGCG treatment (p<0.05).
In addition, we examined the expression of env mRNA in cells treated with 10 µg/mL EGCG for 24 to 96 h (Fig. 3). The env mRNA was scarcely expressed in the uninfected DF-1 cells, whereas the ALV-J-infected cells had higher env mRNA expression at these time points. As we expected, the env mRNA expression decreased significantly by treated with 10 µg/mL EGCG at 24, 48 and 72 h compared to the untreated cells (p<0.05).
The expression of env mRNA in DF-1 cells with different treatments after 24 to 96 h incubation. The error bars represent the SEM. Differences were considered highly significant when p<0.05 (*). The data are representative of the results of three independent experiments.
The effect of EGCG on NF-κB DNA-binding activity
The Trans AM™ NF-κB Family Transcription Factor Assay Kit was used to measure theNF-κB DNA-binding activity in nuclear and cytoplasmic protein extracts from ALV-J-infected cells treated with 10 µg/mL EGCG for 96 h. Among the five NF-kB subunits (p50, p52, p65, c-Rel and RelB), only p50 and p65 significantly translocated from the cytoplasm to nucleus after the cells received the exogenous stimulation (Fig. 4A and B). Under normal growth conditions, p50 and p65 activities were higher in the cytoplasm. When the cells were infected with ALV-J, p50 and p65 translocated from the cytoplasm to the nucleus, but 10 µg/mL EGCG treatment reduced their DNA-binding activity in DF-1 cells.
We also used western blot analysis to determine the protein expression of NF-κB p50 and p65 in both the cytoplasm and nucleus to confirm our result (Fig. 4C). We found that the expression of NF-κB p50 and p65 was higher in the cytoplasm of untreated DF-1 cells. ALV-J infection induced an obvious translocation from the cytoplasm to the nucleus, which was suppressed by the 10 µg/mL EGCG treatment.
The effect of EGCG on NF-kB DNA-binding activity, p50 (A) and p65 (B), and protein expression of NF-kB p50 and p65 in cytoplasm and nucleus (C). The error bars represent the SEM. Differences were considered highly significant when p<0.05 (*). The data are representative of the results of three independent experiments.
DISCUSSION
ALV was discovered more than a century ago and was the first example of an oncogenic virus. It can be passaged by vertical transmission by the virus shedding into the egg albumin or yolk from an infected hen, and by horizontal transmission by ingesting contaminated feces from a viremic hen (Dai et al., 2016). ALV-J is the most harmful pathogenic type, as it damages the immune system and induces immune suppression, resulting in low antibody titers. To date, the use of vaccines to control the disease has not been successful. Moreover, ALV-J-infected chickens are more sensitive to other pathogens, such as Newcastle disease virus, avian influenza virus and reticuloendotheliosis virus (Davidson & Borenshtain 2002, Guo et al., 2010, Dong et al., 2014). Hence, prevention is currently the best approach against ALV-J. Over the last several decades, numerous chemopreventive and/or chemotherapeutic compounds have been identified from natural products including food and plant sources. Out of the natural compounds, polyphenols, especially EGCG in green tea, have gained much attention as potential chemopreventive agents (Jeong et al., 2004). Because ALV is detectable in DF-1 cells earlier than in chicken embryo fibroblasts, DF-1 cells are often used to isolate and detect exogenous ALV (Maas et al., 2006). Therefore, we investigated the protection effect of EGCG on DF-1 cells infected with ALV-J. The results may provide guidance for future in-vivo experiments.
In this study, the cell viability assay demonstrated that at high concentrations (20 and 40 µg/mL) EGCG had a time- and concentration-dependent inhibitory effect on the cell growth of DF-1 cells. Therefore, although EGCG may have a protective effect on ALV-J-infected cells, its concentration should be carefully controlled to avoid growth inhibition in normal DF-1 cells. Previous studies have shown a dose-dependent effect of natural chemopreventive products on DF-1 cell growth. Zhang et al. (2013) have suggested that the safe dose of anthocyanin in blueberries for normal cell growth was less than 80 µg/mL (Zhang et al., 2013). Wang et al. (2014) have found that anthocyanins from purple corn promoted the growth of DF-1 cells in a dose-dependent manner; however, 20 µg/mL anthocyanins obviously decreased cell viability (Wang et al., 2014).
In recent years, EGCG has gained much attention because of its cancer chemopreventive properties, showing remarkable effects in many in-vitro and in-vivo tumor model systems (Nagle et al., 2006). Furthermore, EGCG induced apoptosis in cancer cells without affecting normal cells at physiologically attainable concentrations (Chen et al., 1998). Herein, we examined the protective effect of EGCG on ALV-J-infected DF-1 cells under the safe concentrations. The MTT assay results showed that EGCG treatment of ALV-J-infected cells had a dose-dependent effect, as the lower concentration (5 µg/mL) of EGCG offered no protection to ALV-J-infected cells, whereas the higher concentration (10 mg/mL) did. A similar dose-response was found with EGCG treatment of human malignant tumors, such as epidermoid carcinoma cells and prostate carcinoma cells, which have been reported in a series of articles (Ahmad et al., 1997, Gupta et al., 2000, Sanjay et al., 2003).
Previous research has suggested that the human T-cell leukemia virus type 1 (HTLV-1) transactivator can induce immortalization of primary human T-lymphocytes through a mechanism independent of NF-κB activation. Interestingly, ALV and HTLV-1 belong to the same genus (Wang et al. 2014). Thus, we speculated that the NF-κB pathway may be involved in the response to ALV-J infection. Hence, we assessed the involvement of NF-κB in the EGCG protective effect on ALV-J-infected DF-1 cells.
The NF-κB pathway is involved in inflammation, cancer cell proliferation, invasion, and metastasis (Hoesel & Schmid 2013). In the cell, the transcription factor, NF-κB, is present in the cytoplasm in a complex with its inhibitory protein, IκBa. This complex is primarily inactive; upon activation, IκBa undergoes phosphorylation and degradation through a ubiquitin-dependent pathway, thereby allowing the translocation of NF-κB into the nucleus, to activate various genes responsible for a variety of physiological functions (Ahmad et al., 2000). Activation of the NF-kB pathway is closely linked to an increase in the growth promoting potential and anti-apoptotic responses of cells to a broad spectrum of stimuli, which may result in malignant transformation of cells (Syed et al., 2006). In the present study, we found that NF-κB members were involved in the anti-ALV-J EGCG effect on DF-1 cells. The NF-κB family members, p50, p52, p65 (RelA), RelB and c-Rel, showed higher expression in the cytoplasm than in the nucleus, but the heterodimer of p50 and p65 transferred into nucleus in response toALV-J infection, which resulted in apoptosis. ALV-induced activation of NF-kB, leading to the activation and nuclear translocation of the NF-κB p50/p65, was largely abolished by EGCG treatment, resulting in increased the cell viability. These results suggest that EGCG has the potential to suppress ALV-J-induced cell apoptosis. Our results are consistent with a study by Li et al. (2015) using the human nasopharyngeal cancer cell lines, CNE2 and C666-1, reporting that the active subunit of NF-κB p65, was activated and translocated from the cytoplasm of adherent parent cells into the nucleus; however, a decrease in nuclear p65 localization was observed when these cells were treated with EGCG for 48 h (Li et al., 2015a).
CONCLUSION
In summary, EGCG treatment inhibited ALV-J-induced apoptosis in DF-1 cells in a dose-dependent manner via the NF-κB signaling pathway, and 10 µg/mL EGCG is the optimal concentration. Because ALV-J can induce serious neoplastic diseases, further in-vivo antiviral tests are needed to elucidate whether EGCG can reduce the incidence of tumors via inhibition of viral infection in chicken.
ACKNOWLEDGMENTS
This work was supported by The Application Development Project of Science and Technology Committee in Chongqing (cstc2014yykfB80004), the Foundation and Frontier Project of Science and Technology Committee in Chongqing (cstc2016jcyjA0132), and the Fund of Agriculture Development in Chongqing (The Deeply Utilization of Tea Resource and Products Development).
The authors thank “Edanz Editing China” for revising the English writing.
REFERENCES
- Ahmad N, Feyes DK, Agarwal R, Mukhtar H, Nieminen AL. Green tea constituent epigallocatechin-3-gallate and induction of apoptosis and cell cycle arrest in human carcinoma cells. Journal of the National Cancer Institute 1997;89(24):1881-1886.
- Ahmad N, Gupta S, Mukhtar H. Green tea polyphenol epigallocatechin-3-gallate differentially modulates nuclear factor kappaB in cancer cells versus normal cells. Archives of biochemistry and biophysics 2000;376(2):338-346.
- Chen ZP, Schell JB, Ho CT, Chen KY. Green tea epigallocatechin gallate shows a pronounced growth inhibitory effect on cancerous cells but not on their normal counterparts.Cancer Letter1998;129(2):173-179.
- Dai M, Wu S, Feng M, Feng S, Sun C, Bai D, et al. Recombinant chicken interferon-alpha inhibits the replication of exogenous avian leukosis virus (ALV) in DF-1 cells.Molecular Immunology 2016;76:62-69.
- Davidson I, Borenshtain R. The feather tips of commercial chickens are a favorable source of DNA for the amplification of Marek's disease virus and avian leukosis virus, subgroup J. Avian Pathology 2002;31(3):237-240.
- Dong X, Ju S, Zhao P, Li Y, Meng F, Sun P, Cui Z. Synergetic effects of subgroup J avian leukosis virus and reticuloendotheliosis virus co-infection on growth retardation and immunosuppression in SPF chickens. Veterinary Microbiology 2014;172(3-4):425-431.
- Fan Z, Hu X, Zhang Y, Yu C, Qian K, Qin A. Proteomics of DF-1 cells infected with avian leukosis virus subgroup J. Virus Research 2012;167(2):314-321.
- Gao Y, Liu Y, Guan X, Li X, Yun B, Qi X, et al. Differential expression of immune-related cytokine genes in response to J group avian leukosis virus infection in vivo. Molecular Immunology 2015;64(1):106-111.
- Gao YL, Qin LT, Pan W, Wang YQ, Le Qi X, Gao HL, et al. Avian leukosis virus subgroup J in layer chickens, China. Emerging Infectious Diseases 2010;16(10):1637-1638.
- Guo HJ, LI HM, Cheng ZQ, Liu JZ, Cui ZZ. Influence of REV and ALV-J Co-infection on immunologic function of T lymphocytes and histopathology in broiler chickens. Journal of Integrative Agriculture 2010;9(11):1667-1676
- Gupta S, Ahmad N, Nieminen AL, Mukhtar H. Growth inhibition, cell-cycle dysregulation, and induction of apoptosis by green tea constituent (-)-epigallocatechin-3-gallate in androgen-sensitive and androgen-insensitive human prostate carcinoma cells.Toxicology & Applied Pharmacology 2000;164(1):82-90.
- Gupta S, Hastak K, Afaq F, Ahmad N, Mukhtar H. Essential role of caspases in epigallocatechin-3-gallate-mediated inhibition of nuclear factor kappa B and induction of apoptosis. Oncogene 2004;23(14):2507-2522.
- Hoesel B, Schmid JA. The complexity of NF-κB signaling in inflammation and cancer. Molecular Cancer 2013;12(1):1-15.
- Jeong WS, Kim IW, Hu R, Kong AN. Modulatory properties of various natural chemopreventive agents on the activation of NF-kappaB signaling pathway. Pharmaceutical Research 2004;21(4):661-670.
- Kim HJ, Kim JC, Min JS, Kim MJ, Kim JA, Kor MH, et al. Aqueous extract of Tribulus terrestris Linn induces cell growth arrest and apoptosis by down-regulating NF-κB signaling in liver cancer cells. Journal of Ethnopharmacology 2011;136(1):197-203.
- Lecumberri E, Dupertuis YM, Miralbell R, Pichard C. Green tea polyphenol epigallocatechin-3-gallate (EGCG) as adjuvant in cancer therapy. Clinical Nutrition 2013;32(6):894-903.
- Li YJ, Wu SL, Lu SM, Chen F, Guo Y, Gan SM, et al. (-)-Epigallocatechin-3-gallate inhibits nasopharyngeal cancer stem cell self-renewal and migration and reverses the epithelial-mesenchymal transition via NF-κB p65 inactivation. Tumor Biology 2015a;36(4):2747-2761.
- Li Z, Chen B, Feng M, Ouyang H, Zheng M, Ye Q, et al. MicroRNA-23b promotes avian leukosis virus subgroup J (ALV-J) replication by targeting IRF1. Scientific Reports 2015b;5:10294.
- Liu D, Perkins JT, Hennig B. EGCG prevents PCB-126-induced endothelial cell inflammation via epigenetic modifications of NF-kappaB target genes in human endothelial cells.The Journal of Nutritional Biochemistry 2016;28:164-170.
- Maas R, Van ZD, Oei H, Claassen I. Replacement of primary chicken embryonic fibroblasts (CEF) by the DF-1 cell line for detection of avian leucosis viruses.~Biologicals 2006;34(3):177-181.
- Maruyama W, Shirakawa K, Matsui H, Matsumoto T, Yamazaki H, Sarca AD, et al. Classical NF-kappaB pathway is responsible for APOBEC3B expression in cancer cells.Biochemical and Biophysical Research Communications 2016;478(3):1466-1471.
- Nagle DG, Ferreira D, Zhou YD. Epigallocatechin-3-gallate (EGCG): chemical and biomedical perspectives. Phytochemistry 2006;67(17):1849-1855.
- Payne LN, Brown SR, Bumstead N, Howes K, Frazier JA, Thouless ME. A novel subgroup of exogenous avian leukosis virus in chickens.The Journal of General Virology 1991;72:801-807.
- Prasad S, Ravindran J, Aggarwal BB. NF-kappaB and cancer:how intimate is this relationship. Molecular and Cellular Biochemistry 2010;336(1-2):25-37.
- Sanjay G, Tajamul H, Hasan M. Molecular pathway for (-)-epigallocatechin-3-gallate- induced cell cycle arrest and apoptosis of human prostate carcinoma cells. Archives of Biochemistry & Biophysics 2003;410(1):177-185.
- Silva RF, Fadly AM, Taylor SP. Development of a polymerase chain reaction to differentiate avian leukosis virus (ALV) subgroups:detection of an ALV contaminant in commercial Marek's disease vaccines. Avian Diseases 2007;51(3):663-667.
- Singh BN, Shankar S, Srivastava RK. Green tea catechin, epigallocatechin-3-gallate (EGCG): mechanisms, perspectives and clinical applications. Biochemical Pharmacology 2011;82(12):1807-1821.
- Sun YS, Zhao Z, Zhu HP. Hispolon inhibits TPA-induced invasion by reducing MMP-9 expression through the NF-κB signaling pathway in MDA-MB-231 human breast cancer cells. Oncology Letters 2015;10(1):536-542.
- Syed DN, Afaq F, Kweon MH, Hadi N, Bhatia N, Spiegelman VS, et al. Green tea polyphenol EGCG suppresses cigarette smoke condensate-induced NF-κB activation in normal human bronchial epithelial cells. Oncogene 2006;26(5):673-682.
- Van Aller GS, Carson JD, Tang W, Peng H, Zhao L, Copeland RA, et al. Epigallocatechin gallate (EGCG), a major component of green tea, is a dual phosphoinositide-3-kinase/mTOR inhibitor. Biochemical and Biophysical Research Communications 2011;406(2):194-199.
- Wang D, Lei Y, Ma Y, Zhang L, Zhao X. Inhibition of ALV-A-induced apoptosis in DF-1 cells via inactivation of nuclear transcription factor κB by anthocyanins from purple corn (Zea mays L.). Journal of Functional Foods 2014:274-282.
- Yang ZY, Qu Y, Zhang Q, Wei M, Liu CX, Chen XH, et al. Knockdown of metallopanstimulin-1 inhibits NF-κB signaling at different levels:the role of apoptosis induction of gastric cancer cells. International Journal of Cancer 2012;130(12):2761-2770.
- Zhang L, Lei Y, Gai L, Zhao X, Tong J, Wang D, et al. Study on the Effect of Anthocyanin from Vaccinium uliginosum L. resistance avian leukosis virus subgroup A activity. Acta Agriculturae Boreali-Sinica 2013;28(6):174-180.
Publication Dates
-
Publication in this collection
09 May 2019 -
Date of issue
Jan-Mar 2019
History
-
Received
27 May 2018 -
Accepted
16 Oct 2018